









Геномное разнообразие человечества
- Перейти к Биология | Перейти к новостям научных достижений

Полное секвенирование 300 геномов из 142 популяций со всего мира дало возможность исследователям добавить важные фрагменты в мозаику геномного разнообразия населения планеты. Они пересчитали вклад неандертальцев и денисовцев в современный геном в глобальном масштабе, вычислили, как давно разошлись между собой разные народы, оценили степень гетерозиготности в разных регионах. Последняя намного выше в африканских популяциях. А вот скорость накопления мутаций примерно на 5% выше в популяциях неафриканских народов. Наконец, авторы уточнили источник генофонда жителей Австралии и Новой Гвинеи, показав, что они происходят от тех же популяций, что и жители остальной Евразии; предположение об особом южном пути, по которому пришли их предки, не подтвердилось.
Новая статья, опубликованная в журнале Nature большим коллективом под руководством профессора Гарвардской медицинской школы Дэвида Райха (David Reich) с участием профессора РАН Олега Балановского, представляет ландшафт геномного разнообразия человечества (таково было ее первое название, которое авторы затем заменили на более строгое).
За последние пару лет команда Райха поразила научный мир несколькими исследованиями важнейших проблем – и вот очередное, в буквальном смысле глобальное. В данной работе авторы имеют все основания для глобальных оценок генофонда всего человечества по двум причинам. Во-первых, они проанализировали результаты полного секвенирования 300 геномов из 142 популяций со всех континентов, во-вторых, это секвенирование было проведено с высоким покрытием (в среднем 43х), что гарантирует надежность полученных данных.
В составе международного коллектива есть и российские участники из Института общей генетики РАН и Медико-генетического научного центра в Москве, Института цитологии и генетики, Института молекулярной и клеточной биологии, Института археологии и этнографии и Новосибирского университета в Новосибирске, Института биохимии и генетики в Уфе, Алтайского университета в Барнауле. А среди секвенированных геномов есть и российские образцы.
Чтобы получить картину геномного разнообразия человечества, нужно прочитать много геномов: чем больше, тем точнее будет результат. В проект «1000 геномов», на который ссылаются авторы, входят 26 популяций. Но при этом проект ориентирован в большей степени на задачи медицинской генетики, поэтому в нем игнорируются малочисленные популяции, а для оценки генетического разнообразия они тоже важны. Проект, результаты которого представлены в статье, Simons Genome Diversity Project (SGDP), ориентирован именно на оценку глобального генетического разнообразия.
Исследуемые популяции относились к следующим регионам: Западная Евразия (75 образцов), Центральная Азия и Сибирь (27 образцов), Восточная Азия (47 образцов), Южная Азия (49 образцов), Океания (25 образцов), Африка (55 образцов), Америка (22 образца). Авторы подчеркивают, что популяции в этих регионах были выбраны с тем расчетом, чтобы максимально охватить генетическое, лингвистическое и культурное разнообразие. Россия представлена в базе данных проекта 30 образцами, Северная Евразия (Россия и ближнее зарубежье) – 44 образцами.
Многие проводящиеся до сих пор полногеномные исследования ограничивались секвенированием с низким покрытием генома – например 4х – 6х (цифра показывает, сколько раз был прочитан каждый нуклеотид). Например, именно с таким средним покрытием работал проект «1000 геномов». В данной работе авторы поставили высокую планку и применили глубокое секвенирование – от 34х до 83х (в среднем оно составило 43х). Они выявили 34,4 млн SNP маркеров (однонуклеотидный полиморфизм), 2,1 млн участков с инсерциями/делециями, то есть вставками-выпадениями (INDEL), 1,6 млн коротких тандемных повторов (STR).
Кратко остановимся на главных результатах проекта.
След древних видов в современном геноме
Авторы значительно уточнили генетический вклад архаичных видов человека в геном человека современного вида в разных частях света. До сих пор все подсчеты следов неандертальцев и денисовцев строились на немногочисленном материале. Теперь же на новом количественном уровне удалось статистически пересчитать вклад неандертальских и денисовских предков в геномы и нарисовать полную картину их распределения по миру.
 и денисовских (D) предков в геном современных людей в разных частях света. Цветовая шкала ранжирует этот вклад от 0 до 3% (неандертальцы) и от 0 до 0,5% (денисовцы).")
Оценка вклада неандертальских (С) и денисовских (D) предков в геном современных людей в разных частях света. Цветовая шкала ранжирует этот вклад от 0 до 3% (неандертальцы) и от 0 до 0,5% (денисовцы).
Неандертальский предковый вклад (Рис. С) найден во всех современных популяциях за исключением Африки к югу от Сахары, в Восточной и Южной Азии его больше, чем в Западной Евразии и Америке. Денисовский вклад (Рис. D) несут австрало-меланезийцы и в меньшей степени остальные популяции Восточной и Юго-Восточной Азии.
Время расхождения разных народов
С использованием анализа Мultiple sequentially Markovian coalescent (MSMC), который подсчитывает накопившиеся в паре популяций различия в расчете на один нуклеотид, авторы вычислили время расхождения человеческих популяций различных уровней.
Результаты для каждой пары популяций представлены тремя цифрами, которые показывают, как давно 25%, 50% и 75% генофондов этой пары популяций восходили к общему предку. Это время велико для общих предков африканских и неафриканских популяций. Например, для южноафриканских койсанов и французов датировки расхождения составили 82/131/173 тыс. лет назад, для пигмеев мбути и французов – 67/112/171 тыс. лет назад. В то же время и сама Африка оказалась разделена довольно древними генетическими барьерами. Генетики насчитали шесть различных предковых популяций. Так, с популяцией земледельцев йоруба из Южной Африки койсаны разделились около 58/87/100 тыс лет назад, а мбути и йоруба разделились около 32/56/85 тыс. лет назад. Авторы пишут о потоках генов между различными африканскими популяциями: так, пигмеи мбути, очевидно, по мужской линии смешивались с непигмеями западноафриканского происхождения.
Что касается неафриканцев, то расхождение предковых популяций Западной Евразии, Восточной Евразии и Австрало-Меланезии началось 30-50 тыс. лет назад, вскоре после прохождения самой узкой части «бутылочного горлышка» вышедшими из Африки людьми. А в пределах перечисленных регионов популяции расходились в пределах последних 20 тысяч лет. Так, например, время расхождения между якутами и французами составило 15/23/37 тыс. лет назад, между французами и китайцами – 19/28/42 тыс. лет назад, а между якутами и китайцами уже заметно меньше – 7/10/14 тыс. лет назад.
На этом пути исследователей ожидало немало сюрпризов. Во-первых, выяснилось, что загадочная популяция онге – коренные жители Андаманских островов, которые по внешности и по языку ни на кого не похожи, 20-25 тыс. лет назад имели общих предков с популяциями из Индии и Юго-Восточной Азии. Тем не менее, онге достаточно уникальны, так как в течение последних 10 тыс. лет они находились в изоляции, не были связаны потоками генов с материковыми популяциями.
Проверка «южного пути»
Второй сюрприз состоял в том, что коренные жители Новой Гвинеи и Австралии происходят от гомогенной предковой популяции возрастом 20 тыс. лет, которая достаточно глубоко генетически отделена от остальных неафриканцев. До сих пор считали, что предки жителей Новой Гвинеи и Австралии были изолированы друг от друга со времени первого появления современного человека в этих местах. Возможно, пишут авторы, единой популяции способствовал сухопутный мост, соединявший Новую Гвинею и Австралию вплоть до 10 тыс. лет назад.
Это приводит авторов к дискуссии о том, пришли ли современные люди в Австралию, Новую Гвинею и Андаманские острова по особому «южному пути» или у них был единый путь с материковыми евразийцами. После проверки различными методами они приходят к заключению, что популяции Австралии, Новой Гвинеи и Андаманских островов произошли из того же источника, что и остальные неафриканцы. А если южный путь и существовал, то он привел не к ним, а к древним популяциям, к которым принадлежали образцы Усть-Ишим (45 тыс. лет назад) и Оаse (40 тыс. лет назад).
Этот вывод проиллюстрирован на следующем дереве, которое обозначает предполагаемые генетические связи между неандертальцами, денисовцами, верхнепалеолитическими европейцами, популяциями Восточной Азии, Новой Гвинеи, Австралии и Андаманских островов. Авторы получили хорошее соответствие модели и реальных данных, когда включили в модель потоки генов от неандертальцев и денисовцев. До 4% составил неандертальский поток в геном неафриканцев, до 3% составил денисовский поток в геном австралийцев и папуасов Новой Гвинеи.

Современные популяции обозначены голубым цветом, древние образцы — розовым, предковые линии – зеленым. Пунктирные линии обозначают потоки генов, в том числе от древних популяций. Dinka – этническая группа, проживающая в Южном Судане, Ongе – онге, коренные жители Андаманских островов, Papuan – папуа Новой Гвинеи, Kostenki – образцы с верхнепалеолитической стоянки. Ami и Dai – коренные народы Китая.
Предки жителей Австралии и Новой Гвинеи по этой модели происходят из того же источника, что и предки всех популяций Восточной Азии. Особый южный путь не подтвердился.
Африка продемонстрировала высокое разнообразие
По всем изученным популяциям исследователи вычислили степень гетерозиготности – долю в популяции таких генотипов, в которых на парных хромосомах в одном и том же месте гена находятся разные аллельные варианты. И подтвердили, что наивысшая гетерозиготность по аутосомам (неполовым хромосомам), достигается в Африке южнее Сахары. Как видно на рисунке, африканские популяции по этому показателю (по оси абсцисс) отличаются от остальных со значительным отрывом. По оси ординат указано отношение гетерозиготности по Х-хромосоме к гетерозиготности по аутосомным хромосомам, и по этому показателю Африка снова впереди, а посередине между африканскими и прочими популяциями находятся пигмеи мбути. Вероятно, малую гетерозиготность по этому показателю можно связать с дрейфом «женских» генов.
 и отношение Х-хромосомной к аутосомной гетерозиготности (по оси ординат). Различные регионы обозначены цветными значками.")
Гетерозиготность: аутосомная (по оси абсцисс) и отношение Х-хромосомной к аутосомной гетерозиготности (по оси ординат). Различные регионы обозначены цветными значками.
Еще одним предметом изучения была скорость накопления мутаций. Большой массив данных по проекту SGDP дал возможность сравнить ее в разных популяциях. Исследователи нашли, что в неафриканских популяциях скорость накопления мутаций в геноме на 5% выше, чем в африканских.
Сравнение по предкам
Для сравнения популяций по предковым компонентам авторы применили стандартный метод ADMIXTURE (буквально означает «смесь», «смешение»). О том, как этот метод работает можно прочитать на сайте. Его результаты представлены на рисунке.
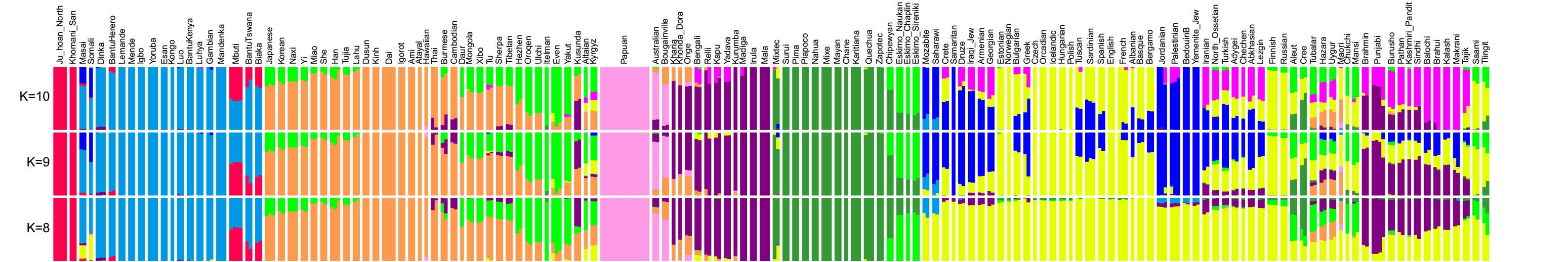
Мы приводим здесь часть спектров, построенных для каждой популяции из расчета, что число их гипотетических предковых популяций (k) равнялось 8,9 и 10. Попробуем прокомментировать результат, полученный авторами для максимального заложенного числа популяций (k=10). Слева мы видим две популяции койсанов (бушменов — красный цвет), затем идут остальные африканские популяции (голубой предковый компонент), из которых выделяются пигмеи, в которых сильно выражен и койсанский, и основной африканский компонент. В популяциях Юго-Восточной Азии преобладают бежевый и ярко-зеленый (сибирский) компоненты. Бледно-сиреневый компонент папуасов Новой Гвинеи в большой степени выражен также у австралийцев и жителей острова Бугенвиль (Соломоновы острова). Фиолетовый цвет обозначает индийский (южноазиатский) компонент; темно-зеленый доминирует у американских индейцев, а у эскимосов индейский компонент сочетается с ярко-зеленым сибирским. Ярко-синий ближневосточный компонент в разной степени представлен у жителей Ближнего Востока, Южной Европы и Кавказа, другой ближневосточный компонент – розовый, охватывает народы Пакистана, окружающих стран и Средней Азии. Лимонный компонент – европейский, он преобладает и у русских, которые по наличию дополнительной небольшой доли сибирского компонента схожи с финнами. У жителей Центральной Азии и Сибири сибирский ярко-зеленый компонент сочетается с европейским и ближневосточным.
Дерево родства народов
Оценка попарных различий между популяциями привели авторов к построению следующего дерева, которое, по их результатам, отражает родственные связи между народами.
. Различными цветами обозначены регионы: Африка (оранжевый), Западная Евразия (розовый), Южная Азия (зеленый), Центральная Азия и Сибирь (голубой), Восточная Азия (голубой), Океания (желтый), Америка (темно-красный).")
Дерево, построенное на основе попарного сравнения популяций (Neighbor-joining tree of relationships based on pairwise divergence statistics). Различными цветами обозначены регионы: Африка (оранжевый), Западная Евразия (розовый), Южная Азия (зеленый), Центральная Азия и Сибирь (голубой), Восточная Азия (голубой), Океания (желтый), Америка (темно-красный).
Изменили ли мутации поведение?
В конце статьи авторы обращаются к гипотезе, что изменения в поведении человека, появившиеся около 50 тыс. лет назад, связаны с одной или несколькими мутациями в генах центральной нервной системы, которые распространились по популяции. Они проверили эту гипотезу и не нашли ее подтверждения. Так что, скорее, к масштабным изменениям в поведении человека привели перестройки не в геноме, а в образе жизни, и культурные инновации.
Метод, использованный для проверки этой гипотезы, состоял в поиске генных локусов, по которым популяции человека современного вида мало отличаются друг от друга (сравнивались койсаны и остальное человечество), но сильно отличаются от архаичных видов (неандертальцев и денисовцев). Ведь именно так должен был бы проявиться отбор по генам в популяции, предковой для всего современного человечества. Среди массы генных локусов, удовлетворявших этому условию хоть в какой-то мере, были отобраны 0,1 % генов (один из тысячи), в которых это условие выполнялось в наибольшей мере, и таких локусов оказалось 38. Для носителей этих генных локусов было определено время жизни их общего предка – по фрагментам хромосом, непосредственно окружающих эти локусы – и среди 38 локусов не нашлось ни одного, для которого большинство пар имеющих его индивидуумов восходили бы к общему предку в последние 100 тысяч лет. А ведь если предполагать, что изменения в поведении 50 тысяч лет назад вызваны новой мутацией, то эта мутация должна была появиться незадолго до этого времени – а все 38 кандидатов оказались старше 100 тысяч лет.
Авторы указывают, что эти результаты не исключают того, что изменения в поведении 50 тысяч лет назад были связаны с генетикой – но исключают то, что генетика была их первопричиной. Ведь 50 тысяч лет назад отбором могли быть подхвачены генетические варианты, возникшие намного раньше и существовавшие в популяции десятки тысячелетий «невостребованными». (Считается, что так произошло с генами, обуславливающими различия в росте северных и южных европейцев (Patterson et al., 2006). Но в данном случае генетические изменения являются не причиной, а следствием: изменившиеся условия жизни создают давление естественного отбора, благоприятствующее определенным внешним признаков, и частоты аллелей, определяющих эти признаки, возрастают. Итак, полученные результаты опровергают теорию, что одна или несколько мутаций определили быстрые изменения в поведении человека за последние 50 тысяч лет.
Результаты показывают, что напротив, изменения в образе жизни благодаря культурным нововведениям были действительной причиной коренных изменений в поведении человека около 50 тысяч лет назад.
Источник: http://генофонд.рф
Latest from Super User
- На ХХ Международной научно-практической конференции «Наступившее будущее: новые форматы, смыслы и сущности образования» обсудили результаты исследований, проведенных научной коллаборацией «Зеленая экономика. Зеленые финансы»
- Член-корреспондент РАН Андрей Наумов: «Спектроскопия — это зрение современной науки»
- «Тихий кризис планеты». Интервью с профессором РАН Еленой Дергачевой
- Шенкарёв Захар Олегович
- Попов Василий Николаевич